Scientific Views/ The Bhaktivedanta Institute
by Richard L. Thompson, Ph.D.

Dr. Richard L. Thompson studied at the State University of New York and Syracuse University and later received a National Science Fellowship. He went on to complete his Ph.D. in mathematics at Cornell, specializing in probability theory and statistical mechanics. His dissertation “Equilibrium States on Thin Energy Shells”—has been published as memoir number 150 of the American Mathematical Society. Dr. Thompson is a disciple of His Divine Grace A.C. Bhaktivedanta Swami Prabhupada and a charter member of the Bhaktivedanta Institute.
One of the most fundamental ideas in modern evolutionary biology is that the physical structures of living organisms can transform from one into another through a series of small modifications, without departing from the realm of potentially useful forms. For example, the foreleg of a lizard can, according to this principle, gradually transform into the wing of a bird, and the lizard’s scales can gradually convert into feathers. In the course of these transformations, each successive stage must serve a useful function for the organism in some possible environment. Thus each intermediate form between leg and wing must be able to act as a serviceable limb under some appropriate circumstances.
Darwin’s theory is based on the hypothesis that, without exception, all the organisms in the world today came about by transformations of this kind, starting with some primitive ancestral form. If such transformations are always possible, then the problem of evolutionary theory is to determine what events in nature might cause them to take place. However, if there exist any significant structures in living organisms that cannot have developed in this way, then for these structures, at least, the hypothesis of evolution is ruled out, and some other explanation of their origin must be sought. Charles Darwin, the founder of the modern theory of evolution, clearly recognized this point: “If it could be demonstrated that any complex organ existed which could not possibly have been formed by numerous, successive, slight modifications, my theory would absolutely break down.” ** (Charles Darwin, The Origin of Species (New York: D. Appleton and Co., 1898), p. 229.)
Although Darwin admitted he could not imagine the intermediate, transitional forms leading to many different organs, he assumed that they might later be revealed by a deeper understanding of the organs’ structure and function, and he proceeded to base his theory on their presumed existence. However, in the nearly 120 years since the publication of his On the Origin of Species, practically no significant advance has been made in the understanding of intermediate forms. While evolutionists often speak of changes in the size and shape of existing organs, they still can do very little but make vague suggestions about the origin of the organs themselves.
The geneticist Richard Goldschmidt once gave a list of seventeen organs and systems of organs for which he could not even conceive of the required transitional forms. These included hair in mammals, feathers in birds, the segmented structure of vertebrates, teeth, the external skeletons and compound eyes of insects, blood circulation, and the organs of balance. ** (Richard Goldschmidt,The Material Basis of Evolution (New Haven: Yale Urtiv. Press, 1940), pp. 6-7.) These organs, and many others, present a fundamental question: How can we explain the origin of a complex system depending on the action of many interdependent parts?
We would like to suggest here that for many organs, the reason why the required chains of useful intermediate forms are unimaginable is simply that they do not exist. Let us try to visualize this in mathematical terms. The class of all possible forms made from organic chemicals can be thought of as a multidimensional space in which each point corresponds to a particular form. We propose that in this space the potentially useful structures will appear as isolated islands surrounded by a, vast ocean of disjointed forms that could not be useful in any circumstances. Within these islands some freedom of movement will exist, corresponding to simple variations in characteristics such as size and shape. But going from island to island-that is, evolving from one particular type of useful organ to another-will require a long and accurate jump across the ocean.
Leaping the Gap
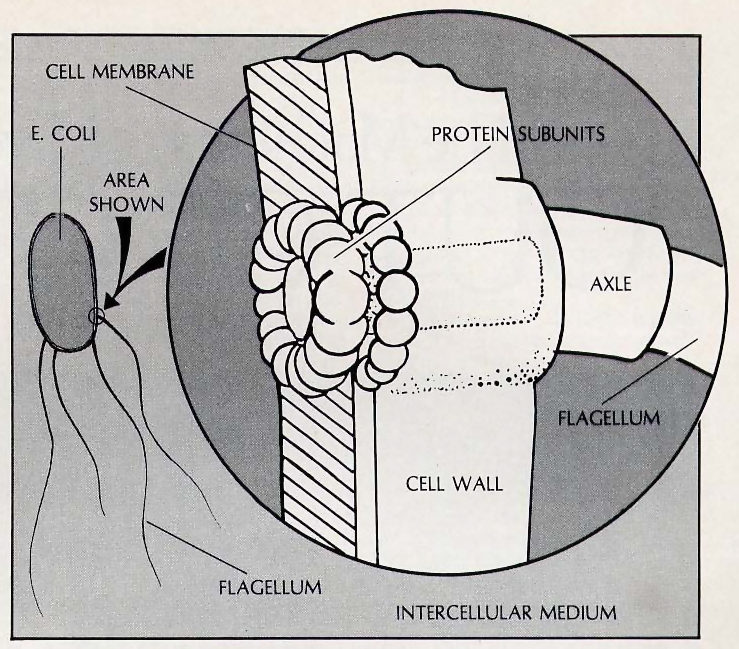
Figure 1 on the previous page depicts these ideas by a mechanical example. Here we consider the space of all possible combinations of mechanical parts, such as shafts, levers, and gears. These mechanical parts are comparable to the molecules making up the organs in the bodies of living beings. Since mechanical parts and molecules alike fit together in very limited and specific ways, a study of mechanical combinations should throw some light on the nature of organic forms.
If we visualize the space of mechanical forms, we can see that some regions in this space will correspond to wristwatches and other familiar devices, and some regions will correspond to machines that are unfamiliar, but that might function usefully in some situation. However, the space will consist mostly of combinations of parts that are useful as paperweights at best.
Since a machine can operate smoothly only if many variables are simultaneously adjusted within precise limits, the useful machines will occupy isolated islands, surrounded by an ocean of machines that are either jammed or broken. If we started from a point on the shore representing a very rudimentary machine, or no machine at all, then we would have to leap over this vast ocean in order to reach, say, a functional wristwatch. As we were making this leap, we could not obtain any guidance by testing the relative usefulness of the forms beneath, for all of them would be equally useless.
Since the bodies of living organisms are built of molecular components and are very complicated, they are not nearly as easy to visualize as the machines in our illustration. However, there are examples of organs that are simple enough to be comparable to man-made mechanisms. One such example is found in the one-celled bacterium Escherichia coli.
Each Escherichia coli cell possesses several long, curved fibers (called flagella) that enable it to swim. ** (Howard C. Berg, “How Bacteria Swim,” Scientific American (Vol. 233, No. 2, 1975), pp. 36-44.) Each flagellum is connected at one end to a kind of motor built into the bacterial cell wall, and when these motors rotate in a certain direction, the flagella rotate in unison and act as propellers to drive the bacterium forward through the water. When the motors rotate in the opposite direction, the flagella separate and change the orientation of the bacterium by pulling in various ways. By systematically alternating between these two modes of operation, the bacterium is able to swim from undesirable to desirable regions of its environment.
The motors are presently thought to be driven by a flux of protons flowing into the cell. Each motor is thought to consist of a ring of sixteen protein molecules attached to an axle, along with a stationary ring of sixteen proteins built into the cell wall. ** (Peter C. Hinkel and Richard E. McCarty, “How Cells Make ATP,” Scientific American (Vol. 238, No. 3, 1978), p. 116.) Protons are steadily pumped out of the cell by its normal metabolic processes. As some of these protons flow back into the cell through the pairs of rings, they impart a rotary motion to the movable ring. Since the motor can operate in forward or reverse, there must be some mechanism that adjusts the molecules in the rings so as to reverse the direction of rotation.
Although the exact details of the Escherichia coli’s molecular motors have not been worked out, we can see that they depend on the precise and simultaneous adjustment of many variables. In the space of possible molecular structures, the functional motors will I represent a tiny, isolated island.
To have a continuum of useful forms spanning the gap between “no motor” and “motor,” we would have to postulate useful organs that do not function as motors but are very similar to motors in structure. For the selective processes of evolutionary theory to eventually choose a working motor, these non-motors would have to be progressively more useful to the bacterium the more motorlike they became. Apart from this very unlikely possibility, evolutionists can suggest no guiding process that can cross the gap.
In the case of very simple organs, such as the bacterial motor, it should be possible to carry out a completely rigorous study of the possibilities of form. Such a study would definitely resolve the question of whether the intermediate forms required by the theory of evolution do or do not exist. Of course, for the highly complicated organs of higher plants and animals, this kind of study may not be practical, but there are still many cases where the combinatorial logic of an organ strongly suggests the impossibility of useful intermediate forms.
One interesting example of this impossibility is found in the statocyst of a certain species of shrimp. ** (W. von Buddenbrock, The Senses (Ann Arbor: Univ. of Michigan Press, 1958), pp. 138-141.) The statocyst is a hollow, fluid-filled sphere built into the shrimp’s shell. It is lined with cells bearing pressure-sensitive hairs and contains a small weight. The weight tends to sink and press against the downward portion of the sphere, thus enabling the shrimp to tell up from down. Curiously, the weight is a small grain of sand that the shrimp picks up with its claws and inserts into the statocyst through a small hole in its shell. The shrimp has to do this every time it moults its shell.
Now, the question is this: By what intermediate stages did the arrangement of the shrimp’s statocyst come about? Both the statocyst and the behavioral pattern involved in picking up the grain of sand are quite complex, and neither is of any use without the other. Even if a statocyst evolved with a built-in weight and then lost this feature by a mutation, the appearance of the insertion behavior would require a leap involving the coordination of many variables.
A Personal Avenue of Approach
At this point, let us try to find an alternative explanation of how such a leap might come about. One natural process in which such leaps are commonly seen is the process of human invention. The products of human creativity, from watches to poetic compositions, are generated with the aid of spontaneous insight, which sometimes enables one to
proceed directly to the solution of a problem without groping laboriously through many false attempts. In fact, it is often the case that after experiencing great frustration in a totally futile trial-and-error search, an inventor will see the complete solution to his problem in a sudden flash. One example of this is the experience of the mathematician Carl Gauss in solving a problem that had thwarted his efforts for years: “Like a sudden flash of lightning,” he wrote, “the riddle happened to be solved. I myself cannot say what … connected what I previously knew with what made my success possible.” ** (Jacques Hadamard, The Psychology of invention in the Mathematical Field (Princeton: Princeton Univ. Press, 1949), p. 15.) It is significant that the solution did not exhibit even a hint of a connection with Gauss’s previous attempts. Here again we find a structure-this time a structure of abstract thought-that is not linked by any discernible chain of intermediate, forms to other, existing structures.
If it is the nature of biological form and the forms of human invention to exist as isolated islands in the sea of possible forms, then some causal agency must exist that can select such forms directly. The experience of inventors indicates that this agency lies outside the realm of human consciousness or control, and that it is capable of acting very quickly.
In Bhagavad-gita a unified description is given of an agency that accounts for the origin of both biological form and human creativity. There it is explained that the ultimate cause underlying the world of our perceptions is not a blind, impersonal process, but a primordial, absolute personality—a personality possessing eternal form, qualities, and activities. Thus in Bhagavad-gita Sri Krsna affirms, “I am the father of all living entities” (Bg. 14.4) and also “I am seated in everyone’s heart, and from Me come remembrance, knowledge, and forgetfulness” (Bg. 15.15).
Of course, even though the nonexistence of intermediate biological forms implies some kind of absolute information or guidance that transcends the categories of ordinary science, this is not sufficient in itself to bring us to the conclusion that the transcendental source must be the Supreme Person. However, this hypothesis opens up very interesting opportunities for further scientific investigation. If the information for the manifestations of form and order in this world is existing in a transcendental state, then this information might be directly accessible in some way. And if the transcendental source is indeed the Supreme Person, as described in the Bhagavad-gita, then it is reasonable to expect that a personal avenue of approach is possible.
In fact, such an avenue does exist. It consists of an elaborate scientific method for establishing a personal relationship with the Supreme. This method, called bhakti-yoga, is quite similar to modern science, in that it depends on clearly specified procedures leading to reproducible results. It is experimentally verifiable, for it is based on direct personal experience attainable by anyone who carries out the procedures correctly.
Organization’s Origins
On the other hand, bhakti-yoga differs from modern science in its method of acquiring basic information. In modern science the hypotheses to be tested, as well as the methods for testing them, are obtained in a haphazard way from the poorly understood sources of “inspiration” or “creative imagination.” In the science of bhakti-yoga, experimental procedures and philosophical principles are both explicitly obtained from the Supreme Person. In other words, although the source of knowledge in both modern science and bhakti-yoga is the Supreme Person, in bhakti-yoga this is fully recognized, and thus there is direct access to the transcendental knowledge available from this source. A good example of this direct access is Bhagavad-gita itself, which, far from being a product of gradual cultural evolution, was directly spoken by Sri Krsna some five thousand years ago at Kuruksetra, in India.
It would be worthwhile for scientists to consider this direct method of attaining knowledge. Even though history has shown that “revealed knowledge” may become corrupted, the basic principle is still valid, and fruitful scientific investigation in this area should be possible. The value of seeking such a rigorous approach is especially apparent if, as we have seen, there is reason to suppose that organized form in both the biological and cultural spheres must originate from a transcendental source.
Leave a Reply