Evolution
A Doctrine in Search of a Theory
Problem:
If the standard evolutionary mechanism is inadequate,
how can the origin of species be explained?
By Sadaputa Dasa

“All reputable evolutionary biologists now agree that the evolution of life is directed by the process of natural selection, and by nothing else.” ** (S. Tax and C. Callender, eds., Evolution After Darwin, Vol. III, Issues in Evolution (Chicago: University of Chicago Press, 1960), pp. 265-266.) With these words Sir Julian Huxley summed up the consensus of learned opinion at the Darwin Centennial Celebration in 1959.
Among the eminent biologists and evolutionists attending the celebration, great confidence prevailed that the origin of living species was now almost fully understood. Evolutionists had clearly established that all living organisms had gradually evolved through small variations in form and function, slowly accumulating, generation by generation, over a vast span of geological time. Geneticists had shown that all biological variations arose from random genetic accidents called mutations. Evolutionary theorists, building on this finding, had clearly identified Darwinian natural selection as the sole guiding force that sorted out these variations and thereby molded the diverse forms of living beings. Although many minute details certainly remained to be worked out, scientists believed they had arrived at an essentially complete understanding of life and its historical development.
With this striking unanimity of established scientific opinion reached little more than two decades ago, perhaps we are surprised to hear that the theory of evolution has recently become the focus of a great controversy among evolutionists themselves. The last few years have seen the established theory of mutation and natural selection increasingly challenged by critical studies and dissenting interpretations of the evidence. The theory has clearly shown itself unsound, although scientists have thus far been unable to devise an acceptable new theory to replace it.
A few months ago this controversy became a near battle as some 150 prominent evolutionists gathered at Chicago’s Field Museum of Natural History to thrash out various conflicting hypotheses about the nature of evolution. After four days of heated discussions (closed to all but a few outside observers), the evolutionists remained convinced that evolution is a fact. Unfortunately, however, they could not reach a clear understanding of just what this fact is. The New York Times reported that the assembled scientists were unable either to specify the mechanism of evolution or to agree on “how anyone could establish with some certainty that it happened one way and not another.” ** (B. Rensberger “Recent Studies Spark Revolution in Interpretation of Evolution,” The New York Times (November 4, 1980), p. C3.)
Why this shift from unanimity and certainty to controversy and indecision? In this article we shall try to answer this question by examining some basic features of the modern theory of evolution. We shall try to identify the reasons why many scientists have sought an evolutionary explanation of life, and we shall also point out some of the problems that have impeded their efforts. We shall argue that the theory of evolution has been motivated more by philosophical misunderstanding than by the strength of empirical evidence and that the current confusion among evolutionary theorists has come about because factual evidence has persistently refused to conform to the patterns imposed by an inconsistent and inadequate philosophical system. Finally, we shall present for these philosophical problems a solution that can lead to a more satisfactory understanding of the nature and origin of life.
Evolution Is Invisible
When Charles Darwin originally set forth his evolutionary theory, he maintained that the forms of living organisms change slowly and continuously from generation to generation and that over many millions of years these changes bring about new species and higher categories of organisms. One immediate implication of this theory was that the fossil record of ancient plant and animal life should display a continuum of fossilized life forms ranging from the most primitive to the most advanced. Given that organisms tend to leave occasional fossilized remains, scientists naturally expected to find a petrified motion picture of evolutionary history entombed in the earth’s sedimentary rocks.
But in Darwin’s time it was well known that the fossil record did not actually reveal such a picture. On the contrary, paleontologists had observed that distinct plant and animal species tended to appear abruptly in the fossil strata without recognizable antecedents. Each species remained essentially unchanged throughout the strata bearing its fossilized remains. Fossils yielded practically no evidence of gradual change from one species to another.
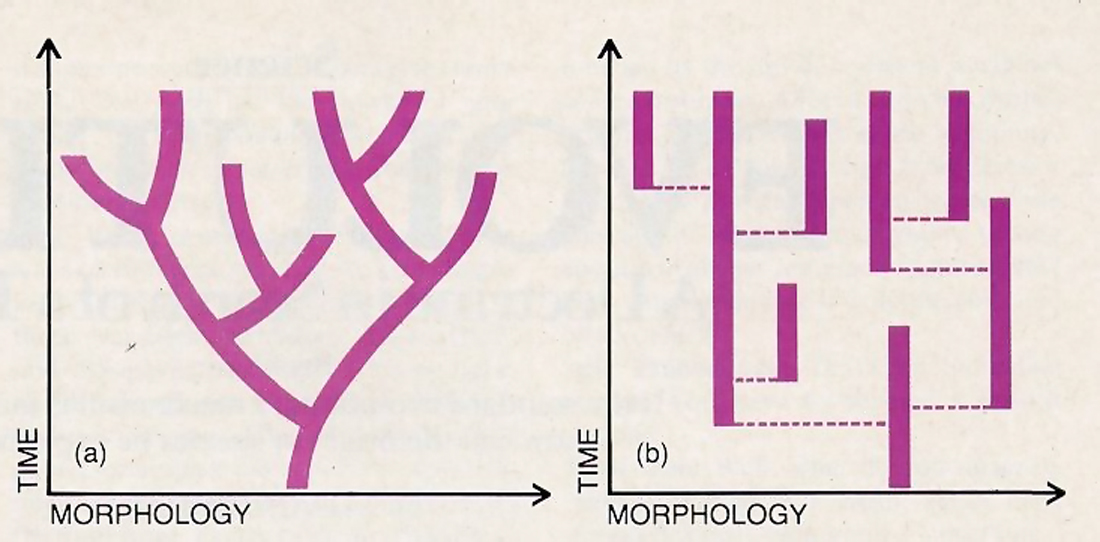
Darwin admitted that the fossil evidence, far from supporting his theory, seemed directly to contradict it. He responded by proposing that the fossil record was drastically incomplete. The innumerable intermediate life forms required by his theory must have existed, but they had left no recognizable traces in the fossil deposits known in his time. Darwin suggested that further research would undoubtedly uncover many of these missing forms, and their discovery would vindicate his theory.
For many years orthodox evolutionary opinion has adhered to Darwin’s basic views. But dissenting voices have increasingly been heard. At the recent meeting of evolutionists in Chicago, Niles Eldridge, a paleontologist from the American Museum of Natural History in New York, declared, “The pattern we were told to find for the last 120 years does not exist.” ** (Ibid.) Despite intense effort, several generations of paleontologists have found few examples in which one fossil species seems to transform gradually into another, and some researchers say none at all have been found.
Punctuated Equilibrium
As a result, Eldridge, Gould, and several other prominent paleontologists now propose that species have not actually arisen by a slow process of transformation As an alternative, they have devised what they call the theory of “punctuated equilibrium.” ** (S.J. Gould and N. Eldridge, “Punctuated Equilibria The Tempo and Mode of Evolution Reconsidered,” Paleobiology, Vol. 3(1977) pp.115-151.) According to this theory, evolutionary changes take place in short bursts separated by long periods during which the forms of living organisms remain static. A typical species will arise from an earlier species in a “geological microsecond”—a period of a few thousand years that appears like an instant from the multimillion-year perspective of geological time. Also, a species will not arise through a gradual modification of its parent population. Rather, it will arise when a tiny group that has been isolated from the main population, perhaps by a geographical barrier, is rapidly transformed.
One consequence of the theory of punctuated equilibrium is that it makes the process of large-scale evolution officially invisible. On one hand, we cannot expect the fossil record to show how a new species evolved, for the evolution takes place in a tiny population during a geological “microsecond.” On the other hand, we cannot expect to see a new species evolve within the recorded span of human history, for a geological microsecond of 10,000 to 50,000 years is still immensely long when measured in human lifetimes.
Of course, we may possibly observe small-scale changes in organisms, like those produced through controlled breeding, or like the famous change in color exhibited by the peppered moths of industrial England. Yet such changes are known to be reversible, and at most they result in only minor variations within a species. For example, settlers introduced domesticated rabbits into Australia in 1788, and some escaped and flourished in the wild. Yet despite the effects of breeding by humans, these domesticated rabbits were still classed as rabbits, and today their descendants have reverted to their ancestral form: they look exactly like wild rabbits. ** (P. Grasse, Evolution of Living Organisms (New York: Academic Press, 1977), p.124.)
But explaining superficial variations of this kind is not the real problem confronting evolutionists. The real problem is explaining how higher forms of plants and animals have arisen from lower forms, and how these in turn have arisen from inanimate matter. No large-scale transformations of this kind have ever been observed within the brief span of human history. The orthodox Darwinian theory maintained that such transformations should be directly visible in the fossil record. But the theory of punctuated equilibrium says we should not expect even the fossil record to show these transformations. In fact, the actual process that brings about new species of life has always been invisible. Now, in the new theory propounded by Eldridge and Gould, this process is held to be invisible even in principle.
The Enigma of Biological Form
If we cannot hope to find direct evidence delineating the development of major new forms of life, we might at least expect the theory of evolution to provide us a convincing explanation of how, in principle, such developments might take place. Since the Darwinian theory asserts that small variations in organic form gradually accumulate, we might expect evolutionary theorists to provide us with plausible evolutionary sequences leading from lower to higher forms of life. In such evolutionary sequences, each organism should be fit to live in its particular environment, and the differences between successive Organisms in the chain should be of the kind we would expect from random mutation.
When we examine the literature of evolutionary theory, we do indeed encounter many explanations of this kind, but in every case they are disappointingly vague and incomplete. Typical is this statement by the prominent evolutionist Ernst Mayr: “The evolution of the eye ultimately hinges on one particular property of certain types of protoplasm—photosensitivity…. Once one admits that the possession of such photosensitivity may have selective value, all else follows by necessity.” ** (E. Mayr, “The Emergence of Evolutionary Novelties,” Evolution After Darwin, Vol. I, The Evolution of Life, S. Tax, ed. (Chicago: University of Chicago Press, 1960), p.359.) Mayr does not, and indeed can not, specify the particular steps leading from a photosensitive speck to a fully developed eye. His account of the evolution of the eye is typical of theoretical evolutionary explanations, for it relies on an abiding faith in the power of natural selection and mutation to effect large-scale transformations in organic form that evolutionists themselves cannot even imagine, much less observe.
Although evolutionists have adhered to this mode of explanation for many years, there is now evidence that its appeal is beginning to wane. According to a report in Science, the predominant view among the evolutionists assembled at the recent meeting in Chicago was that the gradual selective accumulation of small variations cannot account for the appearance of new species. ** (R. Lewin, “Evolutionary Theory Under Fire,” Science, Vol. 210 (November 21, 1980), p.883.)
What has happened is that many evolutionists are now openly acknowledging one of the fundamental problems confronting evolutionary theory—the problem posed by the complex networks of structure and function that are characteristic of living organisms. Generally, each component of such a network is essential for the proper functioning of the whole. How, then, could the complete arrangement have arisen through a finely graded series of functional intermediate forms? In the past many evolutionists have been content to accept on faith that such sequences of intermediate stages must always be possible. But now a number of prominent evolutionists are openly admitting that in many significant cases the required intermediate stages simply may not exist.
To illustrate this problem in evolutionary theory, we shall consider a simple example provided by a type of flatworm called the microstomum ** (W.A. Kepner, W.C. Gregory, and R.J. Porter, “The Manipulation of the Nematocysts of Chlorohydra by Microstomum,” ZoologischerAnseiger, Vol.121 (Jan.-June 1938), pp. 114424.). This flatworm is equipped with a kind of defensive cell called a nematocyst, which can fire a poisonous barbed thread. When the flatworm is attacked by a predator, the nematocysts, situated just beneath the surface of the worm’s back, are discharged, thereby stinging the assailant and driving it away.
The most interesting aspect of this arrangement is that the nematocysts are not produced from the tissues of the flatworm itself. Rather, they are stolen from the hydra, an aquatic organism on which the flatworm preys. The hydra has tentacles armed with several kinds of nematocysts, which it uses to subdue and capture the small animals on which it feeds. Some of these cells fire poisonous barbs, and others discharge various types of coiled and sticky threads that enable the hydra to hold on to its prey.
The flatworms generally avoid hydras. But biologists have observed that when the flatworms need more nematocysts, they will eat hydras and digest all of their tissues except these particular cells. The nematocysts are neither damaged nor discharged, but are enclosed within certain cells, which carry them toward the flatworm’s back. The nematocysts that fire coiled or sticky threads are then digested, but those that fire poisonous barbs are transported to sites just beneath the outer layer of the worm’s back.
There the nematocysts are oriented so that their stings will fire upward. The epithelial cells, which form the worm’s outer layer, become very thin just above the newly positioned nematocysts, thus providing portholes for the firing of the stings. Finally, the cells that have encapsulated the nematocysts undergo extensive changes that enable these cells to act as trigger mechanisms. (The hydra’s original trigger mechanism is contained in a type of cell called a cnidoblast, which the flatworm digests.)
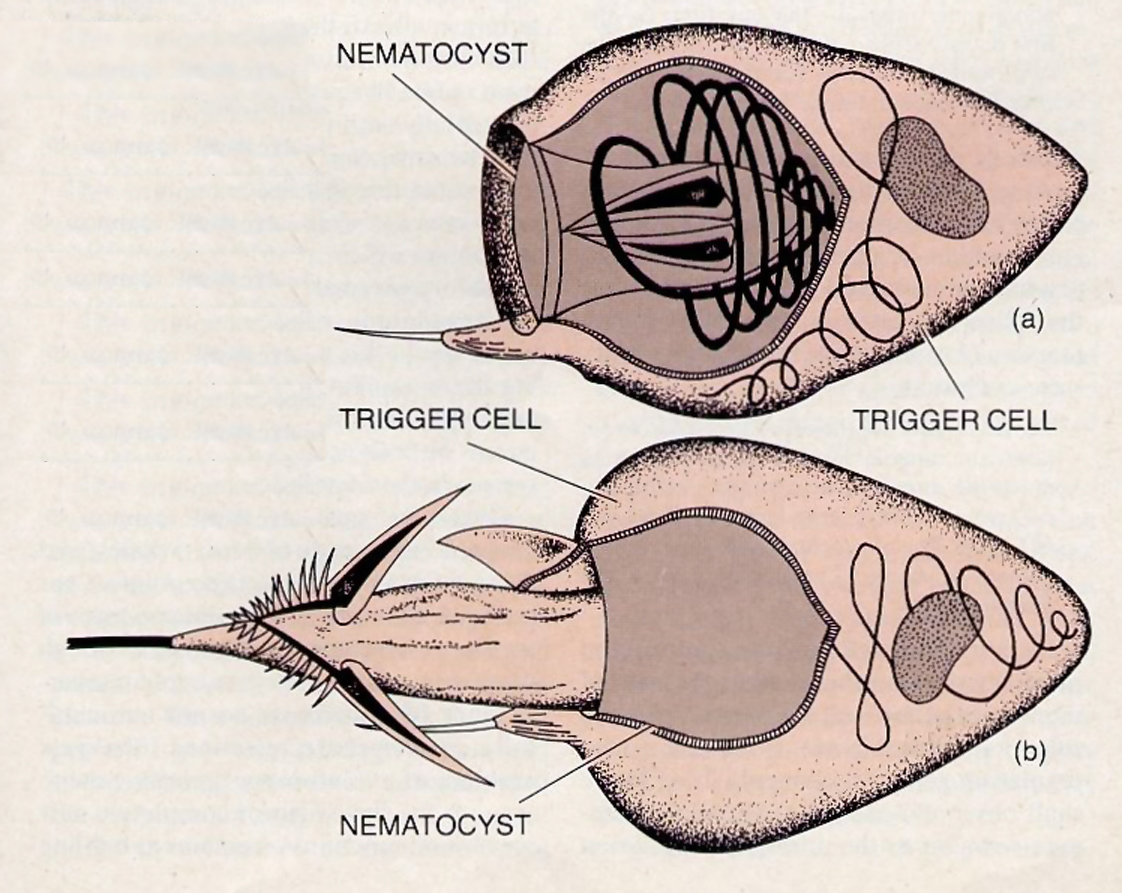
Let us consider whether or not these defensive arrangements of the flatworm could have evolved step by step. An evolutionary scenario would have to begin with an ancestral flatworm that ate hydras but did not make use of their nematocysts.
In such a worm, what would be the first evolutionary step leading to the eventual exploitation of the nematocysts as defensive weapons? Unless the nematocysts were actually used as weapons, for the worm to manipulate them internally would be useless. Indeed, it would be dangerous, since the flatworm can easily be killed by the discharge of the hydra’s stings.
Yet each step in the internal processing of the nematocysts is essential for their eventual use as weapons. If they were not transported to the flatworm’s back, they could not be usefully deployed. If transported to the back but oriented incorrectly, they would be useless or even dangerous. If they were oriented beneath epithelial cells of normal thickness, the discharged sting would lose its momentum while passing through the epithelium, and the worm would sting itself.
There is also a further problem. Evidently the nematocysts are not triggered simply by pressure applied to the worm’s back. Rather, their firing is governed by a complex control mechanism within the worm. Without this trigger mechanism the whole arrangement would be useless, even if the nematocysts were properly oriented beneath epithelial “portholes.”
When examined closely, each step in the internal manipulation of the nematocysts resolves into a complex of substeps. For example, for a nematocyst to be transported to the back of the flatworm, one of the worm’s cells must first recognize it, and then the cell must initiate a process of motion that specifically carries the nematocyst to the worm’s dorsal region. These are both complex procedures. Yet for the flatworm to take advantage of the hydra’s nematocysts, it would seem that many complex arrangements of this kind must be present simultaneously.
We can conclude that the standard Darwinian or neo-Darwinian theory of evolution cannot readily explain the origin of complicated interlocking arrangements such as the flatworm’s defensive system. Yet such systems are by no means rare in nature. Indeed, it might be argued that nearly all complex organs and systems of organs in living beings involve many essential interdependent elements and that they are therefore not amenable to explanation by traditional evolutionary concepts.
The Return Of the Hopeful Monster
We have seen that the standard mechanism of evolutionary theory is inadequate to explain the development of complex living forms. How, then, can the origin of species be explained? At the conference in Chicago there were signs that at least some evolutionists are trying to resurrect a theory that was greeted with almost universal scorn and derision when first proposed in the 1940s. ** (J. Adler, “Is Man a Subtle Accident?” Newsweek (November 3, 1980), pp. 95-96.) This is the theory of “the hopeful monster,” devised by the geneticist Richard Goldschmidt.
The key to this theory is the idea that the genetic systems of organisms must be so arranged that a single mutation can produce, in one stroke, an elaborate systematic change in biological structure and function. Almost all known mutations result in gross defects, and a few result in small modifications that can be useful for the organism under suitable environmental conditions. According to Goldschmidt, however, there must exist a special type of mutation capable of generating new complex structures, such as functional legs, wings, or lungs. Most of these macromutations, as he called them, would result in bizarre monstrosities completely unfit for survival. But a few macromutations would produce “hopeful monsters,” novel creatures that just happened to be adapted to a totally new mode of existence. ** (R. Goldschmidt, The Material Basis of Evolution (New Haven: Yale University Press, 1940).)
This theory is now at a very tentative and speculative stage, and many evolutionists still view it with suspicion. Nonetheless, it represents an important trend in current evolutionary thought, and it illustrates the desperate extremes to which evolutionists have been forced to go in their efforts to construct a workable evolutionary theory. We shall therefore briefly consider some of the reasoning underlying the theory of the hopeful monster.
In its present recension, this theory relies on the concepts of regulative and structural genes.” ** (S.J. Gould, “Hen’s Teeth and Horse’s Toes,” Natural History (July 1980).) Biologists define a structural gene as a sequence of DNA coding that defines a specific structural element of a living organism. An example is the gene for hemoglobin, the oxygen carrying pigment of the red blood cells. In contrast, a regulative gene is a sequence of DNA coding that controls the timing and order of expression of other genes. We can envision an interacting system of regulative and structural genes that acts as a kind of genetic computer program. Such a program might be capable of expressing and repressing various combinations of structural genes in a complex and systematic way. The hopeful monster theory proposes that small changes in such genetic programs might result in the systematic large-scale changes in biological organization that evolutionary theorists need.
Biologists cite certain kinds of mutations as evidence for the existence of regulatory genes. For example, sometimes horses are born with three-toed feet. One might tentatively explain this variation by saying that although the genetic system of the horse always has the structural information for a multitoed foot, in normal horses a regulatory gene suppresses the genes for all the toes but one. When a mutation disables this regulatory gene, the latent genetic information is expressed, and a multitoed horse is born.
A certain mutation of fruit flies provides another possible example of the interaction of regulative and structural genes. In this mutation, known as aristapedia, a fully developed leg grows from the head of the fly in the position where the antenna normally grows. Scientists explain this anomaly by proposing that the regulative and structural genes for the leg comprise a kind of “subroutine” that is set in motion under the control of other regulatory genes. These regulatory genes may store information specifying the location of the leg, and if this information is disrupted by a mutation, the leg may form in an abnormal place.
Although the theory of regulative genes is still highly speculative, it does not seem unreasonable as a way of explaining certain types of mutation. But how this theory can account for Goldschmidt’s hypothetical “macromutations” that produce complex, finely coordinated organs in a single stroke is not at all clear.
Impossible Mutations
We shall try to illustrate the potentialities and limitations of systems of regulatory genes by constructing a simple artificial example. We can regard the following array of symbols as a “genetic” system for a series of English sentences.
I am in (2); the orthodox (1) in (3). I believe I am in (9) as an (8) would be in if set to learn (5). The (8) (1) it was (7); () yet I cannot keep out of (4).
1-would say; 2-thick mud; 3-fetid abominable mud; 4-the question; 5-the first book of Euclid; 6-and tam in much the same mind; 7-no manner of use; 8-old gorilla; 9-much the same frame of mind.
The code used in this genetic system is almost self-explanatory. To produce the encoded English statements, simply replace each number in parentheses with the corresponding numbered phrase. The result is the following statement made by Charles Darwin about the mechanism of large-scale evolution.
I am in thick mud; the orthodox would say in fetid abominable mud. I believe I am in much the same frame of mind as an old gorilla would be in if Set to learn the first book of Euclid. The old gorilla would say it was no manner of use; yet I cannot keep out of the question. ** (Cited in N.C. Gillespie, Charles Darwin and the Problem of Creation (Chicago: University of Chicago Press, 1979), p.87.)
In our artificial genetic system, the numbers in parentheses play the role of regulatory genes, and the phrases play the role of structural genes. If we mutate the regulatory gene (4), changing it to (3), we shall observe a change in Darwin’s statements similar to the aristapedia mutation of fruit flies. (We invite the reader to try this and observe the effects.) Also, when we examine the genetic system closely, we find that a mutation has changed one regulatory gene to (). If we convert this gene to (6), Darwin’s statement apparently acquires an entirely new sentence, although all that has actually happened is that the complete text of the original has been restored.
We can thus see that various kinds of large-scale effects result from mutations in the regulatory genes of our artificial system. Yet all of these effects have one thing in common. They all involve the manipulation of material already present in the genetic system. To induce the system to produce something entirely new is a different matter.
For example, we invite the reader to try to find mutations that will expand Darwin’s remarks to include the following statement from his Origin of Species.
I can see no difficulty in a race of hears being rendered, by natural selection, more and more aquatic in their habits, with larger and larger mouths, till a creature was produced as monstrous as a whale. ** (C. Darwin, On the Origin of Species (London: John Murray, 1959).)
We find ourselves in a quandary. We can gradually build up this additional statement by many small mutations, each of which might occur by “chance” with a reasonably high probability. But the intermediate states will all entail nonsensical sentence fragments that correspond in our analogy to useless or harmful mutant organs; We can also introduce the entire statement by a single random mutation; but then we are confronted by the problem that such a mutation must be exceedingly improbable. The more numerous the letters involved, the more improbable it is that they will fall into place the way you want them. The probability goes down exponentially with the number of variables, and the same can be said in general about the multifeatured biological mutations.
Of course, we could devise a genetic system in which a mutation in a single regulatory gene would cause our new statement suddenly to manifest itself. But could we do this without, in effect, building the statement into the system? If not, one might naturally ask how such complex latent information got into the genetic system in the first place.
Questions such as these cannot be avoided in the study of bears, whales, and living beings in general. The point of our artificial example is that the concepts of regulative and structural genes, although suggesting possible ways to explain several kinds of mutations, do not automatically answer these questions. The real problem of evolutionary theory—how to account for the origin of completely new organs and functions—remains as baffling as before. And until evolutionists can provide a convincing solution to this problem, we must conclude that their evolutionary speculations have no sound basis.
Once again we stress the importance of the problem faced by evolutionary theory: Evolutionists have traditionally declared that mutation and natural selection can fully account for the origin of species, yet suitable sequences of mutations leading to the formation of new organs and systems of organs have never been shown to exist. Until they are, it is pointless even to discuss natural selection, and the origin of species remains shrouded in utter mystery.
Evolution and Negative Theology

We have seen that there is no direct evidence for the evolution of complex organic forms and that some prominent paleontologists maintain that such evidence may never be found. We have also seen that evolutionists do not have an adequate theory of evolutionary change and that they are still groping for such a theory in the realms of speculation and vague conjecture. We are therefore led to ask, In the absence of both observation and theory, what has convinced scientists to accept what we can only call the doctrine of evolution?
One important line of reasoning that has led many persons to adopt an evolutionary point of view could be called the argument by negative theology. Darwin himself used this argument extensively, and since his time it has been a mainstay of evolutionary thought.
In a recent popular book, paleontologist Steven J. Gould sums up the negative theological argument in these words:
“Odd arrangements and funny solutions are the proof of evolution—paths that a sensible God would never tread but that a natural process, constrained by history, follows perforce.” ** (S.J. Gould, The Panda ‘s Thumb (New York: W.W. Norton & Co., 1980), pp. 2O~21.) The basic form of the argument can be outlined as follows: “God must have certain characteristics, and He would have created a certain sort of world. Since the world as we see it is very different from this, it must be that there is no God. Since the only alternative to divine creation that we can think of is evolution, life must have arisen by some kind of evolutionary process.”
This argument breaks down into two basic parts. One of these is the traditional argument from evil against the existence of God. According to this argument, the existence of many kinds of suffering, both in the human species and in the plant and animal kingdoms, is inconsistent with the idea that the world was created by an all-powerful benevolent being. In contrast, such suffering seems to fit naturally into the evolutionary world view.
The second part of the argument is that many features of living organisms would not, as Gould says, be designed by a “sensible God” and must therefore be due to evolution. The extra toes that sometimes appear on the feet of horses are an example of such a feature. Evolutionists argue that God would surely not produce such aberrations but that they can be explained by the hypothesis that horses evolved from a many-toed ancestor.
Another example in support of this argument is provided by Darwin’s work with orchids. Darwin observed that the petals of these flowers are deployed in many remarkable arrangements, which insure that visiting insects will carry pollen from one flower to another. Yet since modified petals, rather than a completely novel kind of structure, are used in these arrangements, Darwin argued that divine creation was ruled out and that the orchids must therefore be products of evolution. In the words of Gould, “If God had designed a beautiful machine to reflect his wisdom and power, surely he would not have used a collection of parts generally fashioned for other purposes.” ** (Ibid.)
What can we say about these arguments? We can observe immediately that they grow from a very much limited and stereotyped understanding of God that is never clearly formulated and that never draws on any specific source of spiritual knowledge. As such, these arguments are certainly unscientific, and when we consider the importance of the theory they are used to support, we can conclude that they are thoroughly irresponsible.
When we consider such negative theological arguments together with the observational and theoretical weaknesses of evolutionary thought, the “theory” of evolution seems little more than a poorly reasoned intellectual reaction against a spiritual tradition that was perceived as inadequate. Moreover, it is an entirely futile reaction, for it has succeeded neither in providing a genuine alternative source of spiritual knowledge nor in establishing a workable material explanation of the origin of life.
Natural Theology: A Dead End
The role the negative theological argument plays in the theory of evolution becomes clear when we consider the historical context in which this theory arose. When Darwin published his Origin of Species in 1859, European thought had been dominated for many years by an approach to spiritual knowledge known as natural theology. According to this approach, one can deduce from observations of natural phenomena that the world has been created by a supremely intelligent, benevolent, and all-powerful being. Pointing to the highly organized structural plans of living beings, the proponents of natural theology maintained that these plans imply the existence of an intelligent creator.
Before Darwin’s time the British philosopher David Hume had pointed out that no amount of finite observation of the things of this world could possibly justify conclusions about an infinite transcendental being. He and many other critics also stressed that the standard conceptions of the nature of this being were inconsistent with some of the most obvious features of the world, for why indeed would a benevolent being create a world of suffering and death? To many scientists, however, the most disheartening aspect of natural theology was that it provided no really satisfying explanation of the origin of life, and it offered no new avenues of exploration. Thus many perceived natural theology as a dead end.
In this context we can understand the initial appeal of Darwin’s theory of evolution. Darwin explicitly rejected the sterile conceptions of natural theology and introduced an approach to the origin of life through entirely physical principles. This approach seemed to bring the question of origins into the familiar realms of physics and chemistry, where scientists had had so much success using the experimental method, and thus it held forth the promise of similar success.
Yet we have seen that this promise has never been fulfilled. Although the theory of evolution has become institutionalized as the standard explanation for the origin of species, it has not out grown its original status as a reaction against an imperfect and restrictive system of theological thought. As such, it is no more valid than the system of natural theology it historically displaced. If speculative reasoning has given rise to an imperfect conception of God, then we certainly cannot expect negative arguments based on this conception to yield a true understanding of life’s origins.
How, then, can we obtain a genuine understanding? Here we shall suggest that we can reach such an understanding only through a valid spiritual science. Speculation resting on a finite set of material observations is indeed inadequate to provide valid knowledge about a supreme transcendental being. But the answer to this problem is not to deny the existence of such a being and to seek explanations solely in familiar physical principles. This is the fallacy of the drunk who lost his keys near the doorstep of his house but would search for them only under a streetlamp because the light was better there.
A Broader Perspective
Here we shall introduce an alternative approach to spiritual knowledge, one that is able to provide a satisfactory understanding of the nature and origin of life. This approach is the spiritual science of sanatana-dharma expounded in the Vedic literatures of India, such as Bhagavad-gita and Srimad-Bhagavatam. As we shall briefly indicate, the unique advantage of sanatana-dharma is that it offers practical procedures that bring the individual into direct contact with transcendence. Sanatana-dharma agrees with the conclusion of natural theology that the world has been produced by a supremely intelligent being. But since sanatana-dharma actually has the kind of sound observational basis that the evolutionists had hoped to attain, it is able to provide a genuine understanding of both the Supreme Being and the relation between the Supreme Being and the material world.
The Bhagavad-gita identifies the ultimate, absolute cause underlying all phenomena as an eternal, transcendental person. This person is known in the Vedic tradition by many names, such as Krsna and Govinda, and is also known as Allah in Islam and as God in the Judeo-Christian tradition. Krsna is the celebrated Supreme Being who is the center of worship in many different religions, and who is also the object of speculative inquiry in diverse schools of philosophy.
According to sanatana-dharma, Krsna is fully endowed with all the essential characteristics of personality, such as mind, senses, emotions, and intelligence. These personal qualities cannot be reduced to transformations of interactions of some impersonal substrate. Sanatana-dharma accepts transcendental personality as the irreducible basis of reality, and all phenomena, whether personal or impersonal, as manifestations of the energy of the Supreme Person.
The concept of personality makes little sense when divorced from the idea of interpersonal relationships. Thus, sanatana-dharma teaches that the Supreme Person is not alone. Krsna is always accompanied by innumerable sentient beings called atmas, who share His inherent personal qualities. These are all eternal, irreducible individuals, but they are qualitatively one with Krsna, and their constitutional nature is to reciprocate with Him in loving personal relationships.
Modern science generally regards concepts such as these as subjects for either blind faith or unverifiable speculation. But in sanatana-dharma the concept of the eternal constitutional nature of the atma provides the key to a practical method whereby the individual person can obtain direct knowledge of transcendental subject matter.
According to sanatana-dharma, each living being in the material world consists of an atma—in association with a body composed of material elements. The atma is bound to this temporary body through false ego, or the illusion that the body is the self. When the atma is in this materially conditioned state, his natural spiritual senses are linked with the sensory apparatus of the body, and he sees himself as a temporary, physical being.
Thus embodied, the atma is oblivious of his original relationship with Krsna and thus unaware of his true potential as a spiritual being. But, according to sanatana-dharma, this is a temporary stare of affairs. Sanatana-dharma provides a practical method, known as bhakti-yoga, whereby the atma can revive his spiritual senses and reestablish direct contact with Krsna. Thus the key to attaining reliable transcendental knowledge is to recognize that each individual person is actually a spiritual being and that his powers of observation are not limited to the sensory machinery of the material body.
The Role of Faith

The subject of bhakti-yoga involves many detailed considerations that lie beyond the scope of this article. Here we shall simply conclude by making a few observations about the role of faith in sanatana-dharma, in modern science, and in the theory of evolution. Since sanatana-dharma is based on verifiable observations, it does not depend on either blind faith or speculative argumentation. But faith is required in any difficult undertaking, and sanatana-dharma is no exception.
For example, before studying modern chemistry the prospective student must have faith that the many experiments upon which the subject is based actually work. He cannot know this in advance, and without such faith he would not be motivated to carry out the arduous work needed to master the subject. Normally, the student will begin with a certain amount of initial faith, and this faith will grow as he acquires more and more practical experience. The same process works in sanatana-dharma.
This brings us back to the theory of evolution. We have observed that this view of life first became prominent as a response to a loss of faith in a particular spiritual tradition. In the beginning of this article we briefly indicated that evolutionists possess neither a substantial body of direct observational evidence nor a workable theory of evolutionary transformation. Thus the theory’s appeal still rests on the power of the negative theological arguments that formed its original basis-arguments that writers on evolution still repeatedly emphasize.
We suggest that these negative arguments appear insubstantial when carefully examined in the light of sanatana-dharma. The traditional “argument from evil” is directly answered by the understanding given in sanatana-dharma of the transcendental nature of the atma and the position of the conditioned atma in the material world. Most of the other negative theological arguments rest on flimsy reasoning and can hold up only until faced by a reasonable alternative.
We have noted that evolutionists cite certain similarities between the bodily parts of various organisms as evidence of a common ancestor. Yet we can also readily, Interpret these similarities as evidence of a common designer. Furthermore, there is no reason to suppose that a “sensible God” would not take advantage of expedient engineering techniques. It might well be quite sensible for God to produce a one-toed horse’s hoof by working from a basic multitoed plan and throwing genetic switches to suppress the development of the other toes. One can deny this possibility only in the absence of a sensible conception of God and a valid understanding of the purpose of the material world.
According to sanatana-dharma, one can understand the purpose of the material creation through the concept of free will. The natural relationship between the atma and Krsna is one of loving service, and love can exist only for one who has freedom. The atma is free to turn away from his relationship with the Supreme Person and seek to be independent, and Krsna creates the material world as a place where the atma can do this. Here the atma becomes temporarily forgetful of his true nature and transmigrates from body to body in various species of life. Therefore, the material world is certainly a place of suffering. The conditioned living beings, deprived of their central object of devotion, inevitably have clashing interests and become sources of intense misery for one another. But should the Supreme Person be held at fault for allowing the living beings to have free will? The evolutionists’ insistence on the “argument from evil” has led nowhere-and it has certainly not alleviated any of the suffering of this world. In sanatana-dharma, however, a genuine solution to the problem of suffering is available. Each individual person, by reawakening his original relationship with Krsna, can remove the root cause of his suffering and attain both unlimited happiness and substantial knowledge about the nature and origin of life.
SADAPUTA DASA studied at the State University of New York and Syracuse University and later received a National Science Fellowship. He went on to complete his Ph.D. in mathematics at Cornell, specializing in probability theory and statistical mechanics.
Leave a Reply